Fotosyntes är den process i vilken växter utnyttjar ljusenergi för att tillverka energirika sockermolekyler från koldioxid och vatten. Det är lätt att tro att mer ljus alltid är bättre eftersom det, intuitivt, borde resultera i mer fotosyntes. Så är dock inte alltid fallet.
Klorofyll är växternas ljusabsorberande pigment och finns i organeller som kallas kloroplaster i alla växtceller som utför fotosyntes. Ljus från solen har en mycket varierande mängd energi (se bild 1). Ljus med hög energinivå (kort våglängd) kan vara så energirik att elektroner i pigmentmolekylerna slås loss från sina elektronskal vilket fragmenterar och förstör pigmentmolekylen. Samtidigt krävs en viss lägstanivå av energi för att ljuset ska vara användbart. Ljus som har väldigt låg energinivå (lång våglängd) kommer, om det absorberas, endast att ge upphov till värme utan att någon energi kan utnyttjas för biokemiska ändamål, vilket inte är fördelaktigt. Naturens lösning är att ljus med för hög eller för låg energinivå helt enkelt inte absorberas. Dessa fotoner passerar rakt igenom växtcellerna eller reflekteras. Endast ljus med en energinivå inom ett för växten lagom intervall, kommer att absorberas. Det ljusspektrum som växelverkar med växter (där energinnivån är lagom) kallas PAR (photosynthetically active radiation) och är snarlikt det spektrum av våglängder som det mänskliga ögat kan uppfatta (ca 380 till 750 nm). Räknas andra ljuspigment in (utöver klorofyll, t.ex. karotenoider och flavonoider) kan växternas spektrum breddas till ca 300 till 750 nm).
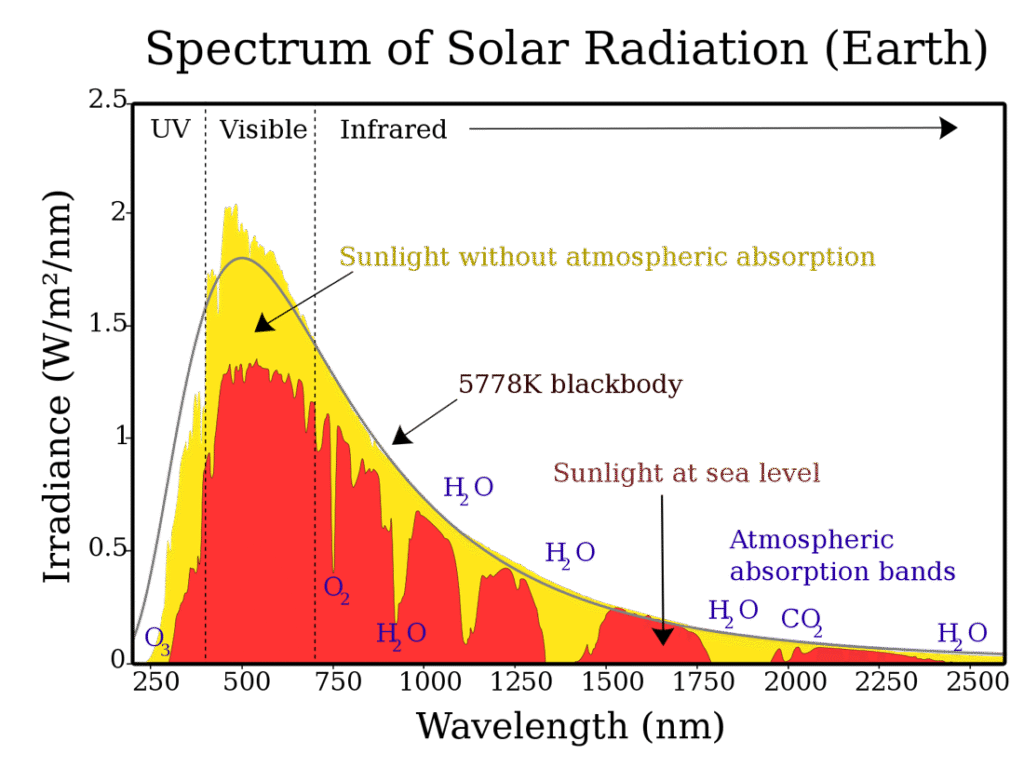
Men även det ljus som absorberas kan komma att bli för mycket för den fotosyntetiserande växtcellen. När en foton väl absorberats av ett pigment så har den exciterat en elektron och tillfört energi (Stark-Einsteins lag) och detta energirika tillstånd måste hanteras av växtcellen. Det optimala sättet är att låta energin användas i fotosyntesen för att konstruera energirika sockermolekyler men denna process är avhängig tillgänglighet av andra faktorer som vatten och koldioxid. Perfekt balans mellan dessa faktorer råder sällan. Exciterade elektroner är mycket instabila och reaktiva vilket kan leda till oönskade och destruktiva reaktioner i cellen. Ett eventuellt överskott på ljusenergi måste kunna hanteras på ett eller annat sätt för att inte riskera skadliga effekter. I växternas fotosyntetiska vävnad finns förvisso andra pigment (t.ex. karotenoider och flavonoider som nämndes ovan) som kan fungera som ett ljusskydd men för den ljusenergi som väl absorberats av klorofyll är det viktigt för cellen att kunna hantera överskottet. Absorberat ljus vars tillförda energi inte förbrukas kan i någon mening liknas vid en trafikstockning av exciterade elektroner.
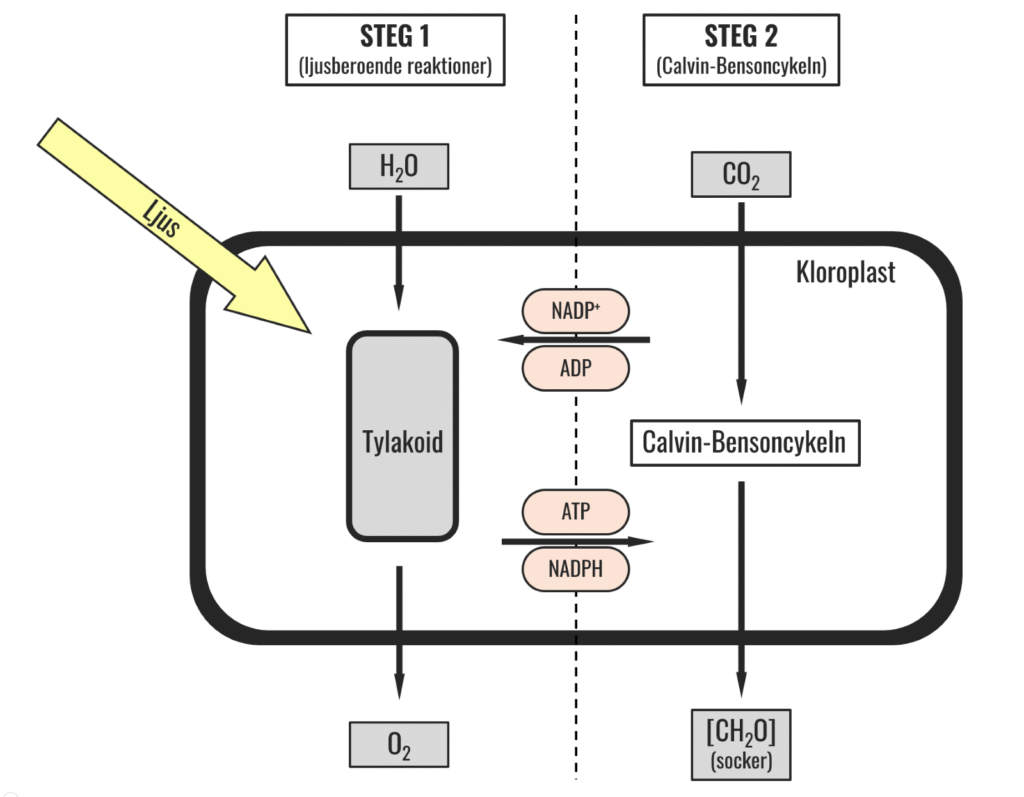
Fotosyntes är en tvåstegsraket (se bild 2). Det första steget (den ljusberoende delen av fotosyntesen) är den som beskrevs ovan där ljusenergi absorberas och i det andra steget (den så kallade Calvin-Bensoncykeln) är målet att fixera kol med hjälp av den energi som utvunnits i steg ett. Med ”fixera” menas att kolatomer, från koldioxid, används i konstruktionen av kolkedjor vilka är stommen i de sockermolekyler som är slutprodukt i fotosyntesen (så kallat fotosyntat, vilket är det begrepp som används som samlingsnamn för alla fotosyntesens produkter). Den första reaktionen då kol ska fixeras katalyseras av ett enzym som går under många namn men som ofta kallas Rubisco (ribulosbisfosfatkarboxylas-oxygenas).
Rubisco använder koldioxid för att utföra en reaktion (där substratet ribulos-1,5-bisfosfat karboxyleras, se bild 3) i vilken slutprodukten är två molekyler av ett trekolsfragment som kallas 3-fosfoglycerat (på engelska förkortas denna molekyl ofta PGA; 3-phosphoglyceric acid). Koncentrationen koldioxid i luft är ca 0,039% eller 390 ppm, och denna låga nivå gör koldioxid till en begränsande faktor i fotosyntesen vid i princip alla ljusnivåer (givet adekvat tillgång på vatten). Ur växtens perspektiv är koldioxid något av en bristvara som växtcellerna måste hushålla med.
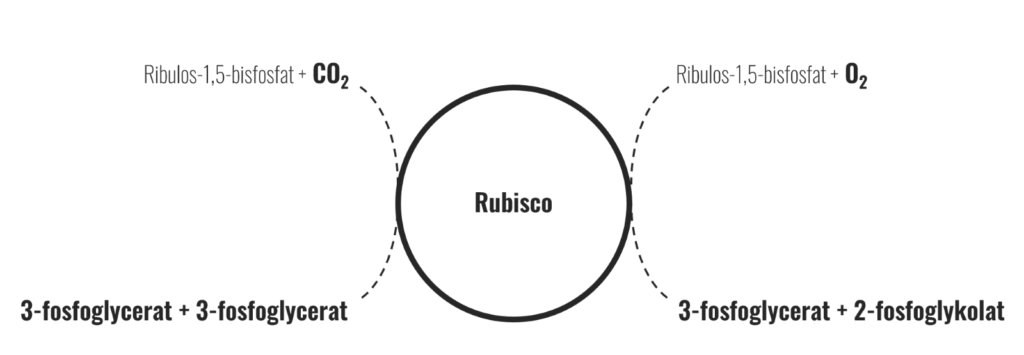
Rubisco kan dock ibland reagera med en syrgasmolekyl i stället för koldioxid (och utför då en annan reaktion där substratet ribulos-1,5-bisfosfat istället oxygeneras). Molekylärt syre finns det mycket av i luft: 20,95% eller 209500 ppm vilket är över 500 gånger högre koncentration jämfört med koldioxid. När Rubisco binder en syrgasmolekyl istället för en koldioxidmolekyl blir resultatet lite annorlunda: i stället för två molekyler 3-fosfoglycerat så bildas endast en och dessutom ett tvåkolsfragment som kallas 2-fosfoglykolat.
2-fosfoglykolat är en problematisk molekyl för växtcellen på två sätt. Dels har den inte något vidare biokemiskt användningsområde vilket gör att värdefullt kol slösas bort. Därutöver är det även toxiskt då det kan inverka och störa andra reaktioner i växtcellen. Det är därför angeläget för växtcellen att göra sig av med 2-fosfoglykolat men samtidigt försöka återvinna dess kolatomer. Denna återvinning kostar energi och kolet kan endast delvis återvinnas. En del av det kol som finns bundet i 2-fosfoglykolat kommer oundvikligen att återgå till koldioxidform och gå förlorat. Effekten är raka motsatsen till vad växten försöker åstadkomma: förlust av kol och förbrukning av energi.
Detta fenomen kallas fotorespiration och ska inte förväxlas med den mitokondriella, oxidativa respirationen (cellandningen). Fotorespiration är en process som slösar bort värdefullt kol och dyrbar energi och det kan tyckas konstigt att ett sådant fenomen tillåts ske i den strikt reglerade växtcellen. Olika teorier har lagts fram till varför inte ett bättre sätt att hantera överskottsenergi har utvecklats. På ett sätt kan man se det som att enzymet Rubisco i princip förväxlar en syrgasmolekyl med en koldioxidmolekyl. En teori är att Rubisco som första enzymatiska steg i Calvin-Bensoncykeln utvecklades under en tid på jorden då syrenivån var mycket lägre och koldioxidnivån var högre. I en sådan miljö skulle förväxling ske väldigt sällan. Det har dock visats att växter som kan utföra fotorespiration i situationer med överskottsenergi från ljus klarar sig bättre än de som inte kan det [1]. Okontrollerad excitation av elektroner kan ge upphov till reaktiva föreningar som ger oönskade reaktioner i cellen vilket potentiellt orsakar permanent skada. Ur växtens perspektiv kan fotorespiration på så sätt ses som ett skydd mot de skadliga effekter som kan uppstå om så många elektroner exciteras att de inte på ett kontrollerat sätt kan göra sig av med den energi som tillförts av ljus. Vid fotorespiration kan växten dumpa sitt energiöverskott genom att leda om alla dessa exciterade elektroner via organeller i cellen som bättre kan hantera reaktiva föreningar (t.ex. peroxisomer och mitokondrier där det finns enzym för att hantera reaktiva föreningar).
Det är önskvärt att i den mån det går undvika de förutsättningar som gör att växten måste utföra fotorespiration. Om växten utsätts för överskottsenergi till följd av överdrivet mycket inkommande ljus så har fotorespiration en skyddande effekt men detta skydd kommer med en förhållandevis stor kostnad för växten. Fotorespiration är, oavsett anledning, en förbrukning av resurser som hade kunnat användas till något bättre, t.ex. tillväxt i rot- eller skottsystem.
Vad kan vi göra för att påverka växtens behov av att utföra fotorespiration?
- Temperatur: det är visat att högre temperatur ökar fotorespiration [2]. Vad som är optimal bladtemperatur varierar mellan arter men för många växter har optimal bladtemperatur visats ligga runt 25°C (något högre för tropiska växter). Vid temperaturer 30°C och uppåt ökar fotorespirationen markant och vid temperaturer över 40°C är fotorespirationen väsentligen lika med fotosyntesen vilket gör att nettoproduktionen av fotosyntat avstannar helt eller t.o.m. blir negativ (om växten utför så mycket fotorespiration att det överstiger fotosyntesens förmåga att kompensera för energiförlusten måste växten förbruka lagrade sockermolekyler och stärkelse).
- Koldioxid: högre koldioxidnivåer minskar fotorespiration. I vanlig utemiljö är det i princip omöjligt att öka koldioxidnivån lokalt runt växterna. Det man däremot kan tänka på är ett bra luftflöde. I luft som är väldigt stilla kan koldioxid i luften som omger bladen, precis i anslutning till bladytan, förbrukas samtidigt som syrenivån ökar till följd av pågående fotosyntes – detta minskar koldioxidnivån och ökar syrenivån lokalt runt bladen vilket ökar sannolikheten att Rubisco binder syre som substrat istället för koldioxid. I inomhusmiljö kan en fläkt som cirkulerar luften upprätthålla goda förutsättningar för fotosyntes. En fläkt kommer även att underlätta för växten när transpirerat vatten ska evaporera från bladen vilket ger bättre temperaturreglering och kan dessutom i någon mån motverka att vissa typer av skadedjur etablerar sig.
- Det har visats att optimal temperatur ökar då koldioxidnivån ökar [3]. För odling i växthus eller växttält där koldioxidnivån kan manipuleras kan den optimala temperaturen ligga över 30°C. Exakt vilken temperatur som är optimal skiljer sig mellan olika arter och med olika koldioxidnivåer.
- Vatten: under torra förhållanden minskar transpirationen för att spara vatten. Transpiration regleras genom gradvis stängning av bladens klyvöppningar (stomata) vilket ger upphov till en proportionerlig ansamling av syre och minskat upptag av koldioxid (vilket ökar sannolikheten för att Rubisco ska binda syre istället för koldioxid). Vi bör således upprätthålla en lagom fuktig miljö i krukan – detta kräver i sin tur vissa förutsättningar avseende bevattning och substrat vilket är ett omfattande diskussionsområde i sig själv. Ett bra substrat är avgörande för att kunna hålla en tillräcklig nivå vatten runt rötterna samtidigt som det ska tillåta adekvat syretillförsel (vilket krävs för rötternas cellandning).
- Salt: salt miljö runt rötterna (i det som kallas rhizosfären, omgivningen runt rötterna) ökar fotorespiration genom att störa det osmotiska upptaget av vatten. Salt miljö kan uppstå över tid om osmotiskt aktiva substanser avsätts i substratet (t.ex. efter gödning). Detta gör att den osmotiska potentialen (ψπ) i rhizosfären blir mer negativ vilket motverkar rötternas vattenupptag. I praktiken har detta samma effekt som torka. Detta är ännu en anledning till att vi bör vattna igenom substratet ordentligt vid varje bevattningstillfälle så att överskott av osmotiskt aktiva föreningar i substratet dräneras.
- Timm S. (2020). The impact of photorespiration on plant primary metabolism through metabolic and redox regulation. Biochemical Society transactions
- Zhang, Z., Zhu, G., & Peng, X. (2024). Photorespiration in plant adaptation to environmental changes. Crop and Environment.
- Cao, W., Tibbitts, T. W., & Wheeler, R. M. (1994). Carbon dioxide interactions with irradiance and temperature in potatoes. Advances in space research : the official journal of the Committee on Space Research (COSPAR)
Lämna ett svar